
Abstract
The objective of the present experimental study was to gain a better understanding of the foraging activity of Asellus aquaticus during fish egg incubation. A. aquaticus were introduced into experimental setups of dead eggs, viable eggs and hatched larvae of zebrafish (Danio rerio), a commonly used model organism. The amount of A. aquaticus and the duration of their exposure to the eggs significantly affected the proportion of consumed dead eggs in each experimental cycle. A. aquaticus belongs to the group of aquatic detritivores, and no predatory behavior was observed during the experiments. These crustaceans could distinguish between the dead eggs and those containing living embryos. Furthermore, zebrafish larvae remained unharmed by A. aquaticus, even in the absence of alternative food source. These findings underscore the potential sanitary role of these crustaceans in natural waters and offer new perspectives on their possible use as biological control organisms in aquaculture hatcheries. Additionally, our results suggest a potential application of A. aquaticus in combating pathogens by reducing the growth substrates for bacteria and fungi.
Similar content being viewed by others


Introduction
The hatching success of fish eggs depends on many factors in nature and in commercial fish producing systems as well. Among many factors, genetic compatibilities, maternal and ova characteristics, spermatozoa motility and sperm/egg ratio play a significant role in fertilization success and the hatching rate of fish embryos1,2,3. Besides the internal biological features, environmental factors such as optimal temperature, salinity, predators, chemical pollutants, and diseases are all crucial factors for successful hatching4,5,6,7.
Bacterial and fungal infections are a primary concern during the incubation period of fish eggs, especially in aquaculture hatcheries8,9,10,11. Harmful pathogens such as Saprolegnia sp., Flavobacterium sp. or Pseudomonas sp. are commonly and naturally found in many water systems. The mucous layer of eggs appears to be a good substrate for adhesion and colonization by many of these pathogens12.
It is generally observed that the appearance of pathogens, such as bacteria and fungi, correlates with the proportion of dead eggs4,13. However, it is possible that increased growth of bacteria and fungi is a result of, not necessarily a cause of, increased mortality. On the other hand, dead eggs may promote the spread of horizontally transmitted diseases. Prior studies have shown Saprolegnia parasitica to initially colonize dead eggs and the hyphae from these then infect the surrounding living embryos14. Smith et al.13 showed that only dead eggs are infected by zoospores, whereas both live and dead eggs were infected by hypheal growth from adjacent infected eggs. In contrast, it has also been observed that S. diclina from fungus-infected salmonids can directly infect live eggs without any need for prior colonization of dead eggs15.
In addition to the potential transmission of diseases between eggs, there could be other adverse effects resulting from a high quantity of unfertilized and deceased eggs within a water system. Dead and ruptured eggs may provide considerable nutrients for heterotrophic bacteria by leaching16. Large numbers of bacteria can have high oxygen requirements17, producing metabolic by-products18 or toxins19. The proliferation of heterotrophic bacteria not only influences egg survival rate but it has negative impacts on a recirculating aquaculture system (RAS) via the competition with nitrifying bacteria20.
The physical barrier of the chorion and membranes defends against invading pathogens21; after hatching, the exposure to pathogens dramatically increases. Colonized dead eggs also create a route for pathogen transmission towards the freshly hatched larvae.
Various techniques such as egg disinfection have been applied to control egg pathogens8,10 in commercial aquaculture systems. Chemical treatments have been used to control disease transmission with varying levels of success. However, the elimination of dead eggs, with their potential as a substrate for disease and infection in fish, has been of limited focus. In a separate study, the mechanical removal of deceased eggs of Chinook salmon (Oncorhynchus tshawytscha) at the eyed stage resulted in a reduction in bacterial counts4. This technique could be applicable to salmonids owing to their relatively larger egg size, enhanced visibility, and the absence of a swirling incubation medium. However, it entails a significant investment of time and labour. For other species (e.g., cyprinids or percids) in which egg size is considerably smaller or incubation occurs in turbulent media or on surfaces22, the manual elimination of dead eggs becomes challenging, ineffective, and at times unfeasible.
Thus, biological control methods can serve as effective alternatives to chemical or labour-demanding mechanical disease management. The concept of the biological control of pests is gaining interest as the harmful effects of agrochemicals continues to be understood. Grass carp, Ctenopharyngodon idella, is one of the best-known examples of a biological control “tool” to manage invasive aquatic plants23. Several species of wrasse (Labridae) are used as cleaner fish to remove salmon lice (Lepeophtheirus salmonis) from farmed Atlantic salmon (Salmo salar)24. The first application of macroinvertebrates as a biological control of fungal disease was reported by Oseid25. In this study, it was shown that grazing on the mycelia of oomycetes infected eggs by Asellus militaris and Gammarus pseudolimnaeus increased the hatching rate of walleye embryos (Sander vitreum). However, the application of crustaceans as a biological control in fish hatcheries is still not very common.
A. aquaticus is a native detritivore abundant in most European freshwater ecosystems. High densities can be found in waters with organic pollution or in relatively clean water with high levels of naturally occurring organic matter26. Several studies have concluded that A. aquaticus selectively feed on detritus colonized by microorganisms, in particular fungi27,28. We do not have proof that fish eggs which are not infected by pathogenic fungi attract A. aquaticus. Our hypothesis was that A. aquaticus can consume not just the mycelia from eggs infected with oomycetes, but is also capable of eliminating or reducing the number of dead eggs before the excessive outgrowth and spread of pathogenic fungi, without harming living embryos or freshly hatched larvae. We implemented three experimental cycles, investigating the foraging preferences of A. aquaticus using different ratios of living and dead eggs/embryos and freshly hatched larvae of zebrafish (Danio rerio) and evaluating the effects of A. aquaticus’ presence on larval vitality.
Materials and methods
Ethics statement
According to the European Directive on the protection of animals used for scientific purposes (2010/63/EU) zebrafish embryos/larvae are not subject to ethical regulations until they reach independent feeding stage (120 hpf)29. No independently feeding zebrafish larvae (older than 120 hpf) were involved in our experiments.
Zebrafish maintenance
A genetically unmodified, laboratory zebrafish strain (AB) was used for the experiments. This line has been bred in the Zebrafish Laboratory of the Hungarian University of Agriculture and Life Sciences (HUALS). Fish were maintained in 3 L polycarbonate tanks in a recirculated system (ZebTEC, Tecniplast S.p.a., Italy) through an upwelling bead filter at 25 ± 2 °C, and fed three times per day with commercial flakes (Sparos Zebrafeed, 400–600 µm) and live Artemia nauplii grown from cyst (Ocean Nutrition > 230000NPG). The photoperiod was set to 14 h light: 10 h dark. Guidelines from the good laboratory practice (GLP—Organization for Economic Co-operation and Development (OECD)) and Institutional Animal Care and Use Committees of the Hungarian University of Agriculture and Life Sciences were followed for animal care. The National Food Chain Safety Office, Animal Health and Animal Welfare Directorate of Government Office of Pest County has registered the site of the Zebrafish Laboratory as an animal testing user, breeder, and supplier facility and authorized its operation under the file number PEI/001/1719–2/2015 (Appendix I.).
Broodstock of A. aquaticus
A. aquaticus specimens were collected from an experimental recirculating aquaculture system (RAS) of HUALS designed for fish broodstock housing. Three 5000 L fish tanks were connected and operated as a freshwater RAS which included a drum filter, moving bed bio reactor and aeration. Daily water exchange varied between 2 and 6% of the total volume depending on the biomass kept in the tanks. The system has been in operation for over five years without shut down. For broodstock we offered live zooplankton as supplementation. We assume that A. aquaticus appeared in the system owing to this practice as zooplankton was collected from natural waters. The starting population then multiplied on the organic particles in the effluent water. A. aquaticus was collected from the effluent water with a plastic sieve in a weekly quantity of approximately 200–1500 individuals. Specimens for the experiments were chosen randomly, the only criteria were that adult individuals (4–5 mm) were used.
Propagation, egg collection and incubation prior to the experiment
During propagation, five zebrafish females with five males were introduced into each breeding tank (n = 6). According to the protocol of zebrafish fertilization and embryo isolation (http://www.zfic.org/common%20techniques/mating.pdf), all fish were released to spawn. 1.7 L breeding tanks (ZebTEC, Tecniplast S.p.a., Italy) were used in all experiments. Water conditions were: temperature 25 °C; pH 7.0 ± 0.2; average conductivity 525 µS. Eggs were collected from each tank and placed into Petri dishes (Ø 100 mm) 2 h after the onset of light. Eggs were incubated in an incubator (25.5 °C, photoperiod was set at 14 h light: 10 h dark) with daily water changes. After a 24 h incubation period, all eggs were checked and separated into two batches: dead eggs (white) and living eggs (transparent with living embryos). The number of dead eggs represents both unfertilized eggs and embryos that have died during the first 24 h of embryogenesis.
Incubation of zebrafish embryos in the presence of A. aquaticus: experimental design
The experiments were carried out in sterile, flat-bottom, non-treated, multiwell-plates with a lid (6—well format, Vtotal = 5 mL, Vwater = 4 mL). In the first experiment (E1), A. aquaticus individuals (total body length 4–5 mm) were added to some of the wells in varying numbers (1, 3 or 5 per well; Table 1). Prior to the experiments, A. aquaticus specimens were starved in each well for 24 h. Varying ratios of dead eggs and living embryos were introduced into the wells at 24 h post-fertilization (hpf) (Table 1). Each well contained a piece of biological filtration media (AQ-09KL 436 m2/m3, Ø/ = 9/7 mm, Aquacultur GmBH), which served as a hiding/hanging place for A. aquaticus. In order to provide identical experimental conditions, the control wells without A. aquaticus contained the same media. The number of living embryos and dead eggs were checked every five hours till the end of E1. In the second experiment (E2), dead eggs, freshly hatched larvae (non-feeding stage) and A. aquaticus (total body length 4–5 mm) were introduced into the wells at different ratios (see Table 1). The number of larvae and dead eggs was monitored every ten hours until the end of the experiment. Water temperatures were 27.4–27.8 °C and 24.5–24.7 °C during E1 and E2, respectively. The photoperiod was set at 14 h light: 10 h dark without water changes during the experimental cycles.
Evaluation of larval vitality
To evaluate the effects of A. aquaticus presence on the vitality of zebrafish larvae, experiment 3 (E3) was performed. Normally developed 24 hpf zebrafish embryos were placed in 6-well cell culture plates (5 embryos/well) containing 10 mL system water per well and incubated at 25.5 ± 0.5 °C until 118 hpf in the presence or absence of A. aquaticus (3 individuals/well) and in the presence or absence of dead zebrafish eggs (3 dead eggs/well) (n = 30 larvae/group). We provided one piece of biofilter media per well as shelter for A. aquaticus. The experimental groups were as following: (I) control (living embryos); (II) living embryos + dead eggs; (III) living embryos + A. aquaticus; (IV) living embryos + dead eggs + A. aquaticus (Table 1). Mortality was monitored daily. At 118 hpf, A. aquaticus individuals and biofilter media were carefully removed. At the end of the experiment (E3) morphological and behavioral alterations were detected (see below).
Detection of morphological alterations (E3)
Zebrafish larvae were anesthetized, positioned laterally and imaged under a stereo microscope (Leica M205 FA, Leica DFC7000T camera, Leica Application Suite X software, Leica Microsystems GmbH, Germany). The standard and total body length, the swim bladder area, and the length of the gut were assessed using ImageJ software (n = 30 larvae/group). The approximate gut length was determined based on the work of Chuang et al.30.
Zebrafish behavioral assay (E3)
Zebrafish larvae were individually transferred into 96-well cell culture plates containing 200 µL system water per well (n = 30 larvae/group). Next, the 96-well plates were placed (one by one) in a ViewPoint Behavior Technology instrument and the larvae were monitored during a 10 min light—20 min dark—10 min light incubation period. The ViewPoint ZebraLab system continuously tracked the movement of the larvae and analyzed their locomotor activity (e.g., the total distance covered by the larvae in small or in large movements) at one-minute intervals. To evaluate alterations in the locomotor activity of the larvae, we segmented the behavior test into distinct periods and determined the total distance traveled by the larvae in one minute during each specified period. These periods included: the pre-transition period (5 min before the light–dark transition), light–dark transition (the first one minute of the dark phase) and first and second halves of the dark phase.
Statistical analysis
The effects of time and the number of A. aquaticus was tested in the first experiment (E1) using a Multivariate Analysis of Variance (MANOVA), in which the ’number of eggs containing living embryos’, the ’number of dead eggs’ and the ’number of hatched larvae’ was considered as dependent variables and ’hours’ and the ’number of A. aquaticus’ were factors. In the second (E2) experiment, there was no variation in the number of larvae, therefore a two-way factorial ANOVA was performed, in which ’number of dead eggs’ was considered as the dependent variable, and ’hours’ and the ’number of A. aquaticus’ were factors. Dependent variables were transformed using log(x + 1) to satisfy the assumptions of MANOVA and ANOVA. All analysis were conducted in SPSS v25 statistical software. Data derived from zebrafish vitality tests were analyzed by one-way parametric ANOVA and post-hoc Tukey test or Kruskall-Wallis test and post-hoc Dunn test using the GraphPad Prism 8 software. Results are presented as the mean ± standard deviation (SD).
Results
Effects of A. aquaticus presence on the number of dead eggs, living embryos and hatched larvae (E1)
In the first experiment, the number of dead eggs continuously decreased after the first 5 h in A. aquaticus containing wells. The decreasing proportion of dead eggs accelerated according to the number of A. aquaticus specimens/well. At the first assessment time (5:00 h), the number of eggs were statistically different (p < 0.05) between the experimental groups and this difference increased with time (Tables 2 and 3, Fig. 1). The first larva hatched during the 30th hour in every experimental group, however, the hatching process was more dynamic in the presence of A. aquaticus compared to control.

Effects of A. aquaticus on the number of dead eggs, living embryos and hatching rate during the incubation period (E1). Stacked bar chart presents the average number of dead eggs, living embryos, disappeared eggs and hatched larvae in presence or absence of A. aquaticus over a 45-h incubation period. Initially, each replicate was introduced with 10 dead and 20 living embryos. The bar graphs indicate that a substantial number of the introduced dead eggs disappeared during incubation when A. aquaticus was present.
Effects of A. aquaticus presence on the number of dead eggs and living larvae (E2)
Similar to the previous experiment (E1), the number of dead eggs continuously decreased from the 5th hour in A. aquaticus containing wells. A. aquaticus did not harm the non-feeding larvae and there was no natural mortality detected, even when there was no alternative food source for the crustaceans. All introduced larvae survived throughout the experimental period, irrespective of the number of coexisting A. aquaticus specimens (Table 4, Fig. 2).

Effects of A. aquaticus on larval survival, with or without dead eggs as alternative feed source (E2). The stacked bar chart displays the number of dead eggs, disappeared eggs and hatched, live zebrafish larvae. Bar graphs indicate that the number of introduced hatched, live larvae remained unchanged during the 40-h incubation period, regardless of the presence or absence of dead eggs and/or A. aquaticus.
Effects of A. aquaticus presence on larval vitality (E3)
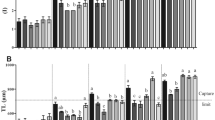
To assess the effects of A. aquaticus presence on larval vitality we determined the morphological and behavioral changes in the 120 hpf zebrafish larvae. There was no mortality during the experiments, although we did observe 10% whole body malformations in the co-presence of dead eggs and A. aquaticus during embryonic development. Regardless of this phenomenon, we did not detect any dramatic morphological alterations in the other larvae (Fig. 3A–E). No differences were found in the standard body length (Fig. 3B) however, interestingly, a slight but statistically significant decrease could be detected in the total body length and the gut length of the larvae developed in the presence of dead eggs compared to control (Fig. 3C,D). The gut length of the larvae developed in the presence of dead eggs also differed significantly from the group containing only A. aquaticus (Fig. 3D). Additionally, there was a more obvious and statistically significant decrease in the swim bladder area after incubation in the presence of dead eggs and in the co-presence of dead eggs and A. aquaticus as compared to the control and to the group containing only A. aquaticus (Fig. 3E).

Effects of A. aquaticus presence during embryonic development on zebrafish larval morphology (E3). (A) Representative images of 120 hpf zebrafish larvae developed in the presence or absence of A. aquaticus and/or dead zebrafish eggs. Standard body length (B), total body length (C), length of the gut (D) and the swim bladder area (E) of 120 hpf zebrafish larvae developed in the presence or absence of A. aquaticus and/or dead zebrafish eggs. Data represent the mean and SD. Different letters indicate statistically significant differences, p < 0.05.
Next, we investigated the locomotor activity of the 120 hpf larvae performing the zebrafish behavioral assay. Locomotor activity measurement in zebrafish embryos and larvae is a frequently used model system for assessing the effects of environmental stressors. Normally, zebrafish larvae exhibit a sudden increase in locomotor activity following a transition from light to dark, followed by a decrease throughout the dark and the subsequent light period. These changes in the locomotor activity are regarded as characteristic behaviors of zebrafish at early larval stages31,32. Consequently, this allows for the comparison of potential differences in larval activity across various test periods. At the end of the incubation, we could not detect significant differences in the response of the larvae to the light–dark transition between the groups (Fig. 4A,C,F,H). However, the larvae which were developed in the presence of dead eggs or in the co-presence of dead eggs and A. aquaticus showed a significant decrease in locomotor activity during the pre-transition period (5 min before light–dark transition) (Fig. 4A,B,F,G), and a significant increase in the first half of the dark phase post transition compared to controls and to the group containing only A. aquaticus (Fig. 4A,D,F,I). Interestingly, the total distance covered by the larvae in small movements showed a slight, but significant decrease during the dark phase in the group containing only A. aquaticus as compared to every other group (Fig. 4A,D,E). However, this phenomenon was not observed for large movements (Fig. 4F,I,J).

Effects of A. aquaticus presence during embryonic development on the locomotor activity of zebrafish larvae (E3). The figure summarizes the behavioral response of 120 hpf larvae developed in the presence or absence of A. aquaticus and/or dead zebrafish eggs during light–dark-light exposure. Under control conditions, the transition from light to dark elicits a sudden increase in the locomotor activity of zebrafish larvae. This activity decreases throughout the dark and the second light period. The whole test phase was divided into different periods to assess the potential alterations in the locomotor activity between the experimental groups. Locomotor activities were expressed as the distance traveled by the larvae with small or large movements. Line graphs showing the total distance covered by the larvae in small (A) and large (F) movements during a 10 min light—20 min dark—10 min light incubation with one minute resolution. Space between dashed lines (denoted as ‘1’, ‘2’, ‘3’, ‘4’) represents the statistically analyzed periods (300–600 s. (B,G) 600–660 s. (C,H) 660–1200 s. (D,I) 1200–1800 (E,J)) of the test. Data represent the mean and SD. Different letters indicate statistically significant differences, p < 0.05.
Taken together, the presence of A. aquaticus during the embryonic development did not result in substantial morphological alterations, however, it slightly affected the locomotor activity of the larvae in a certain phase of a light–dark period. Moreover, dead eggs alone had a more remarkable impact on the measured morphological and behavioral parameters than the presence of A. aquaticus.
Discussion
In the classical approach of biological control, predators and parasitoids are used which target the pathogens or pests themselves. The first attempts at chemical free control of oomycete parasites using two invertebrates, Asellus militaris, and Gammarus pseudolimneaus, were reported by Oseid25. According to their results, both invertebrates improved the survival rate of eggs by preventing fungal growth; however, Gammarus showed some predation on live eggs and larvae. Our knowledge is limited about the presence and significance of this phenomena in natural fish stocks.
The scraping feeding strategy of A. aquaticus makes it possible to ingest mycelia selectively from the surface of dead organic materials28. Bloor33 conducted feeding preference experiments with A. aquaticus and found that they prefer detritus which was partially digested by microbial communities and fungi. Possible reasons behind this experimental result could be that fungi can eliminate allelopathic chemicals of plants and through partial digestion they are able to make detritus more utilizable for the detrivores, which can also use the fungal enzyme system to degrade organic materials. Fungi also enrich the feed with their own micro- and macronutrient content34.
In this study, our theory on how A. aquaticus eliminates mycelia encompasses three main aspects: (a) there is direct grazing on the egg surfaces, aligning with their inherent feeding strategy; (b) we noted the consumption of infected and dead eggs; and (c) as a consequence of the movements and feeding behaviors of A. aquaticus, the eggshell of dead eggs can incur physical damage, potentially leading to “disappeared” eggs, as observed in this investigation. We believe that in this case the chorion undergoes damage, causing organic material to disperse into the water, which impedes the eggs from functioning as a substrate for fungal growth. The results of our study show that the feeding habits of A. aquaticus mitigate infection and prevent fertilized eggs from being entangled in fungal mycelium. (Fig. 5).

Schematic figure of the foraging features of A. aquaticus during fish embryo incubation. Dead and/or infected fish eggs can serve as a food source for A. aquaticus. These crustaceans have the ability to differentiate between dead and/or infected fish eggs and living embryos or larvae.
During our experiments none of the hatched larvae were harmed by A. aquaticus, even if no other feed source was provided. Other researchers found the same, that A. aquaticus specimens, as opposed to other crustaceans like Gammaridae, do not cause harm to young fish larvae25,35. However, it was also observed that hatching occurs faster in the presence of A. aquaticus. Most likely, the cause of this is the abrasion of egg shells due to the movements and feeding activity of the crustaceans. This phenomenon did not correlate with the number of A. aquaticus per well under our experimental conditions. This could be beneficial in a way, as shortened incubation periods also reduce the chance of fungal infection, though shorter incubation often results in smaller, weaker larvae, or limited hatching rate. Although we did not observe significant changes in the morphology of the larvae developed in the presence of A. aquaticus, the co-presence of dead eggs and A. aquaticus resulted in a low percentage (10%) of malformations. Besides, minor alterations were detected in the locomotor activity of the larvae in the presence of dead eggs and/or A. aquaticus. At the end of embryonic development, zebrafish larvae possess a complex and functional sensory system36,37. Consequently, the presence of A. aquaticus during the incubation period could affect the larvae's responses in behavioral tests conducted after incubation. It is important to note that most significant changes in morphology and locomotor activity were detected in groups where dead eggs were present. Detailed characterizations of the effects of dead eggs on the embryonic development of fish species are limited. Nevertheless, it can be hypothesized that the observed morphological and behavioral alterations may result from decaying organic matter. An elevated amount of organic matter can lead to hypoxia and release compounds that may affect the behavior and developmental rate of the embryos38,39,40. However, the exploration of the causal relationships related to these processes exceeds the scope of this study.
In summary, the presence of A. aquaticus during the zebrafish embryonic development had no considerable effects on larval vitality, at least until 120 hpf. However, it is possible that the greater density and larger size of A. aquaticus may have negative effects on the outcome of the incubation of relatively small, vulnerable fish eggs, which calls for further investigation.
Our present study demonstrates that A. aquaticus can reduce the number of dead zebrafish eggs. In addition, our preliminary experiments indicated that A. aquaticus was able to reduce the mass of water mold hyphae colonizing the eggshells of brown trout (Salmo trutta fario), a species with larger eggs (diameter of 5 mm) and longer embryonic development (44 days) as compared to zebrafish, and they were also able to consume the dead trout eggs (Supplementary Fig. 1). The elimination of dead eggs by A. aquaticus in large-scale operations can be limited. Hence, commercial hatcheries might need to administer multiple isopod inoculations to replace the manual egg picking method for disease management. A more feasible application of A. aquaticus could be with the semi-natural spawning of endangered fish species (e.g., Misgurnus fossilis and Carassius Carassius) or high value ornamental fish, where egg incubation and larvae rearing take place in the same unit, and the breeders do not show cleaning and fanning behavior. Control of pathogens by the elimination of the substrate for bacterial and fungal growth could improve the hatching rate and larval survival as well as general fish hygiene.
The applicability of this method in hatcheries should be tested in detail and the conditions of breeding and application of A. aquaticus should be optimized considering the effects of density, mating period, moulting and the different habits of the sexes and life stages34,41.
Conclusions
According to the results of the current study, A. aquaticus can distinguish between living and dead fish eggs and are able to eliminate or reduce the number of dead eggs which can serve as a media for bacterial and fungal growth. We found that A. aquaticus do not harm eggs containing viable embryos and hatched larvae, even when alternative food sources were not available. We suggest that A. aquaticus can be a potential alternative biological control organism in special cases when small-scale egg incubation and larvae rearing are carried out at the same place (e.g., ornamental fish breeding or ex situ conservation). However, further investigations are required to determine the appropriate settings for utilizing this species in biological control, considering the specific fish species and environment involved.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
References
Johnston, T. A. et al. Hatching success of walleye embryos in relation to maternal and ova characteristics. Ecol. Freshw. Fish 16, 295–306 (2007).
Rodríguez-Muñoz, R. & Tregenza, T. Genetic compatibility and hatching success in the sea lamprey (Petromyzon marinus). Biol. Lett. 5, 286–288 (2009).
Gallego, V., Pérez, L., Asturiano, J. F. & Yoshida, M. Relationship between spermatozoa motility parameters, sperm/egg ratio, and fertilization and hatching rates in pufferfish (Takifugu niphobles). Aquaculture 416, 238–243 (2013).
Barnes, M. E., Ewing, D. E., Cordes, R. J. & Young, G. L. Observations on hydrogen peroxide control of Saprolegnia spp. during rainbow trout egg incubation. Prog. Fish. Cult. 60, 67–70 (1998).
St Mary, C. M., Gordon, E. & Hale, R. E. Environmental effects on egg development and hatching success in Jordanella floridae, a species with parental care. J. Fish Biol. 65, 760–768 (2004).
Cook, M. A., Guthrie, K. M., Rust, M. B. & Plesha, P. D. Effects of salinity and temperature during incubation on hatching and development of lingcod Ophiodon elongatus Girard, embryos. Aquac. Res. 36, 1298–1303 (2005).
Mhadhbi, L. & Beiras, R. Acute toxicity of seven selected pesticides (alachlor, atrazine, dieldrin, diuron, pirimiphos-methyl, chlorpyrifos, diazinon) to the marine fish (turbot, Psetta maxima). Water Air Soil Poll. 223, 5917–5930 (2012).
Subasinghe, R. P. & Sommerville, C. Disinfection of Oreochromis mossambicus (Peters) eggs against commonly occurring potentially pathogenic bacteria and fungi under artificial hatchery conditions. Aquac. Res. 16, 121–127 (1985).
Wagner, E. J., Ronney, E. A., Billman, E. J., Forest, A. & Cavender, W. Comparison of the efficacy of iodine, formalin, salt, and hydrogen peroxide for control of external bacteria on rainbow trout eggs. N. Am. J. Aquacult. 70, 118–127 (2008).
El-Dakour, S., Saheb, A. I. & Al-Abdul-Elah, K. Effects of commonly used disinfectants on bacterial load, hatchability and survival of Bluefin Sea bream (Sparidentex hasta) eggs. Aquac. Res. 46, 1281–1291 (2015).
Marking, L. L., Rach, J. J. & Schreier, T. M. American fisheries society evaluation of antifungal agents for fish culture. Prog. Fish Cult. 56, 225–231 (1994).
Vatsos, I. N., Thompson, K. D. & Adams, A. Colonization of rainbow trout, Oncorhynchus mykiss (Walbaum), eggs by Flavobacterium psychrophilum, the causative agent of rainbow trout fry syndrome. J. Fish Dis. 29, 441–444 (2006).
Smith, S. N., Armstrong, R. A., Springate, J. & Barker, G. Infection and colonization of trout eggs by Saprolegniaceae. T. Brit. Mycol. Soc. 85, 719–764 (1985).
Kitancharoen, N., Yuasa, K. & Hatai, K. Effects of pH and temperature on growth of Saprolegnia diclina and S. parasitica isolated from various sources. Mycoscience 37, 385–390 (1996).
Rand, T. G. & Munden, D. Involvement of zoospores of Saprolegnia diclina (Oomycotina: Saprolegniaceae) in the attachment to and invasion of eggs of brook trout under experimental conditions. J. Aquat. Anim. Health 5, 233–239 (1993).
Bell, G. R., Hoskins, G. E. & Hodgkiss, W. Aspects of the characterization, identification, and ecology of the bacterial flora associated with the surface of stream-incubating Pacific salmon (Oncorhynchus) eggs. J. Fish. Res. Board Can. 28, 1511–1525 (1971).
Hansen, G. H. & Olafsen, J. A. Bacterial colonisation of cod (Gadus morhua) and halibut (Hippoglossus hippoglossus) eggs in marine aquaculture. Appl. Environ. Microb. 55, 1435–1446 (1989).
Léonard, N., Guiraud, J. P., Gasset, E., Cailleres, J. P. & Blancheton, J. P. Bacteria and nutrients—Nitrogen and carbon—In a recirculating system for sea bass production. Aquacult. Eng. 26, 111–127 (2002).
Kawai, A., Yoshida, Y. & Kimata, M. Biochemical studies on the bacteria in aquarium with circulating system: 1. Changes of the qualities of breeding water and bacterial population of the aquarium during fish cultivation. B. Jpn. Soc. Fish 30, 55–62 (1964).
Michaud, L., Blancheton, J. P., Bruni, V. & Piedrahita, R. Effect of particulate organic carbon on heterotrophic bacterial populations and nitrification efficiency in biological filters. Aquacult. Eng. 34, 224–233 (2006).
Castro, R. et al. Disparate developmental patterns of immune responses to bacterial and viral infections in fish. Sci. Rep.-UK 5, 1–13 (2015).
Schlumberger, O. & Proteau, J. P. Reproduction of pike-perch (Stizostedion lucioperca) in captivity. J. Appl. Ichthyol. 12, 149–152 (1996).
Treasurer, J. W. A review of potential pathogens of sea lice and the application of cleaner fish in biological control. Pest Manag. Sci. 58, 546–558 (2002).
Skiftesvik, A. B. et al. Wrasse (Labridae) as cleaner fish in salmonid aquaculture –The Hardangerfjord as a case study. Mar. Biol. Res. 10, 289–300 (2014).
Oseid, D. M. Control of fungus growth on fish eggs by Asellus militaris and Gammarus pseudolimnaeus. Trans. Am. Fish. Soc. 106, 192–195 (1977).
Murphy, P. M. & Learner, M. A. The life history and production of Asellus aquaticus (Crustacea: Isopoda) in the River Ely, South Wels. Freshw. Biol. 12, 435–444 (1982).
Rossi, L. & Fano, A. E. Role of fungi in the trophic niche of congeneric detrivorous Asellus aquaticus and A, coxalis (Isopoda). Oikos 32, 380–385 (1979).
Graça, M. A. S., Maltby, L. & Calow, P. Comparative ecology of Gammarus pulex (L.) and Asellus aquaticus (L.) I: Population dynamics and microdistribution. Hydrobiologia 281, 155–162 (1994).
Strähle, U. et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 33, 128–132 (2012).
Chuang, L. S. et al. Zebrafish modeling of intestinal injury, bacterial exposures and medications defines epithelial in vivo responses relevant to human inflammatory bowel disease. Dis. Model Mech. 12(8), dmm037432 (2019).
Du, W., Chen, X., Shi, M., Bian, F. & Zhao, Z. Ethanol affects behavior and HPA axis activity during development in zebrafish larvae. Sci. Rep. 10(1), 21402 (2020).
Hill, B. N. et al. Assessment of larval zebrafish locomotor activity for developmental neurotoxicity screening. In Experimental Neurotoxicology Methods (eds Llorens, J. & Barenys, M.) 327–351 (Springer, 2021).
Bloor, M. C. Dietary preference of Gammarus pulex and Asellus aquaticus during a laboratory breeding programme for ecotoxicological studies. Int. J. Zool. 2011, 1–5 (2011).
Rosenthal, G. A. & Janzen, D. H. Herbivores: Their Interaction with Secondary Plant Metabolites (Academic Press, 1979).
Taylor, N. G. & Dunn, A. M. Size matters: Predation of fish eggs and larvae by native and invasive amphipods. Biol. Invasions 19, 89–107 (2017).
Moorman, S. J. Development of sensory Systems in Zebrafish (Donio rerio). ILAR J. 42(4), 292–298 (2001).
Neuhauss, S. C. Zebrafish vision: Structure and function of the zebrafish visual system. In Fish Physiology Vol. 29 (ed. Neuhauss, S. C.) 81–122 (Academic Press, 2010).
Kalueff, A. V. & Cachat, J. M. (eds) Zebrafish Models in Neurobehavioral Research (Humana Press, 2011).
Shang, E. H. & Wu, R. S. Aquatic hypoxia is a teratogen and affects fish embryonic development. Environ. Sci. Technol. 38(18), 4763–4767 (2004).
Sear, D. A. et al. Does fine sediment source as well as quantity affect salmonid embryo mortality and development?. Sci. Total Environ. 541, 957–968 (2016).
Kinzler, W. & Maier, G. Asymmetry in mutual predation: Possible reason for the replacement of native gammarids by invasives. Arch. Hydrobiol. 157, 473–481 (2003).
Acknowledgements
We would like to express our gratitude to Mr. György Hoitsy (Hoitsy & Rieger Ltd.) for providing the trout eggs, and to Mr. Péter Kiss for his assistance in the laboratory. The authors thank Ms. Izabella Berta for the contribution to the maintenance of the zebrafish broodstock.
Funding
Open access funding provided by Hungarian University of Agriculture and Life Sciences. This work was supported by the National Research Development and Innovation Office of Hungary (NKFI K—135824) and the ÚNKP-23-4 (ÚNKP-23-4-II-MATE-4) and ÚNKP-23-3 (ÚNKP-23-3-I-MATE/22) New National Excellence Program of the Ministry for Culture and Innovation from the source of the National Research, Development and Innovation Fund.
Author information
Authors and Affiliations
Contributions
Conceptualization, Tamás Müller, Quyến Nguyễn Ngọc, Bence Ivánovics, Máté Havasi, Ádám Staszny, Béla Urbányi and Balázs Kucska; Methodology, Tamás Müller, Quyến Nguyễn Ngọc, Máté Havasi, Ádám Staszny, Károly, Vranovics, Béla Urbányi, Bence Ivanovics, Ádám Varga and Balázs Kucska; software, validation, Tamás Müller, Máté Havasi and Balázs Kucska; formal analysis, Tamás Müller, Quyến Nguyễn Ngọc, Ádám Staszny, Béla Urbányi, Bence Ivanovics and Balázs Kucska; investigation, Tamás Müller, Quyến Nguyễn Ngọc, Károly Vranovics, Béla Urbányi, Ádám Varga; resources, Tamás Müller, Béla Urbányi and Balázs Kucska; data curation, Tamás Müller, Ádám Staszny and Béla Urbányi; writing—original draft preparation, Tamás Müller, Bence Ivánovics, Quyến Nguyễn Ngọc, Máté Havasi, Ádám Staszny, Károly, Vranovics, Béla Urbányi, Jeffrey Daniel Griffitts and Balázs Kucska; writing—review and editing, Máté Havasi, Jeffrey Daniel Griffitts; visualization, Bence Ivánovics, Tamás Müller, Máté Havasi and Balázs Kucska; supervision, Máté Havasi; project administration, Quyến Nguyễn Ngọc, Ádám Staszny and Béla Urbányi; funding acquisition, Tamás Müller and Béla Urbányi. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kucska, B., Ngọc, Q.N., Ivánovics, B. et al. Removal of dead fish eggs by Asellus aquaticus as a potential biological control in aquaculture. Sci Rep 14, 7295 (2024). https://doi.org/10.1038/s41598-024-57903-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57903-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
